Apuntes sobre el electrocardiograma en la infancia y la adolescencia
Rev Argent Cardiol 2024;92:309-312. http://dx.doi.org/10.7775/rac.v92.i4.20802
Correspondence: Samuel Sclarovsky. E-mail: samuel_s@netvision.net.il

Physiological hypertrophies in childhood
At birth the mammalian ductus arteriosus closes and pulmonary peripheral resistance is reduced. Right ventricular predominance persists for a limited time. Left ventricular predominance appears in rats at 2 weeks, in pigs at 6 weeks (1) and in humans at 12 weeks. (Figure 1) It is amazing to observe small calves and foals in the meadows, which at 4 hours after birth, can run at 40 - 60 km/h. For this to occur, they must have a left ventricle perfectly adapted to survive. Evolution prepares the left ventricle during pregnancy to support intense efforts a few hours after birth.
The newborn has a heart rate of 190 to 200 bpm and a high rate is maintained during childhood, between 80 and 120 bpm. (2) This heart rate is determined by catecholamines. Adrenaline is secreted by 2 sources, one endocrine (the adrenal gland) and one paracrine (the cardiac terminal sympathetic fibers). In the myocytes there are alpha and beta receptors. Exaggerated stimulation of the beta receptors is very harmful to the myocyte. (3) To avoid this effect there is a biological beta-blocker, arrestin. (4) But the effect on alpha receptors (in the myocyte membrane there are 3 types of alpha receptors) stimulates the formation of sarcomeres. (5) Sarcomeres aggregate longitudinally. (5,6) The effect of adrenaline on the alpha receptor determines the growth of sarcomeres and myocyte elongation, which defines the QRS voltage.
But there is another electromechanical system that stimulates physiological hypertrophies. In the endocardium of the left ventricle there are tension receptors, whose distribution in the subendocardium is not homogeneous. (7) It is possible to speculate that the morphology of the child's electrocardiogram varies, depending on the concentration of tension receptors in the endocardium. Higher concentration of tension receptors, plus stimulation of alpha receptors, will induce physiological hypertrophies.
In 30% of cases the electrocardiogram in children is normal (Figure 2) In 70% of cases, during childhood, there is physiological hypertrophy that accompanies body growth. The ECG can record 4 different phenotypes of hypertrophy: 1) apical (Figure 3), which is expressed with dominant R waves in V4 and V5; 2) anteroseptal (Figure 4), which is expressed with dominant R waves in V2 (upper third of the septum), V2 and V3 (upper and middle third of the septum), and V2 to V4 (upper, middle and lower third of the septum); 3) posteroseptal (Figure 5), which is expressed with deep S waves in V2, V3, and V4; and 4) lateral, which is expressed with high R waves in lead I and deep S waves in AVR and V1. In rodents, inhomogeneously distributed endocardial tension receptors have been identified in the septum and lateral apex. (8) It could be suspected that the inhomogeneous distribution of tension receptors is a factor of universal importance in cardiac function in mammalian childhood.
Fig. 2
6-year-old boy without eccentric physiological hypertrophy, and probably concentric physiological hypertrophy.


Fig. 4
9-year-old boy with physiological eccentric anteroseptal hypertrophy, with high R waves in V2, V3 and V4.


Normal ECG tracing could be due to concentric hypertrophy, homogeneous distribution of tension receptors or a mutation in the hypertrophic cascade. (9) It is important to note that also in 30% of cases adults with systolic overload do not present eccentric hypertrophy (personal experience). Electrical/mechanical activity induces a feedback reaction. Electrical activity stimulates the Z-line and titin protein to increase tension in the actin-myosin complex. (10)
There is a genetic fetal program in embryonic life, which has a mildly hypoxemic environment. (11) In this context mitochondria use glucose to generate energy. But with birth, the mitochondria begin to use oxygenated fatty acids as energy fuel. The fetal program remains hidden, inactive; but in cases of physiological or pathological hypertrophies it is reactivated. This program prevents the progression of hypertrophies towards dilatation and heart failure. We speculate that in physiological cardiac hypertrophies in children a genetic fetal program may exist as myocardial protection. Interestingly, physiological hypertrophies in children are no longer seen in adolescence.
Gender hormones determine the repolarization morphology of the electrocardiogram in adolescence
Estrogens control the fast transient outward rectifier potassium current, (12) which forms the downward limb of the T wave, which is slow and elongated compared with this limb in the male gender, and also the slow rectifier potassium current, (13) which forms the peak of the T wave, which is very low or negative compared with the male high and acuminate T wave.
These hormones also control calcium influx in the phase 2 of the action potential, expressed with an elongated ST segment. (14)
Androgens have a stimulatory effect on potassium (15) and calcium channels, which is expressed by a high and acuminate T-wave peak, a short and fast descending limb, and short and elevated ST segment. (15) The effect of testosterone on the calcium channel influences the ST segment, which is short and elevated usually in V2 and V3. (16) (See Figure 6 y 7)
Fig. 6
ECG of a 22-year-old male.

The red arrow points to the fast rectifier potassium current, which acts on phase 3 of the action potential (AP) and forms the short, fast descending limb of the T wave.
The blue arrow points to the slow rectifier potassium current, which acts at the junction of phases 2 and 3 of the AP and forms the high, acuminate T wave. The green arrow points to the calcium current that acts in phase 2 of the AP and conditions the short and elevated ST segment on the ECG.
From Nerbonne JM, Kass RS. Molecular physiology of cardiac repolarization. Physiol Rev. 2005;85:1205-53. Modified by Sclarovsky S
Fig. 7
ECG of a 16-year-old woman.
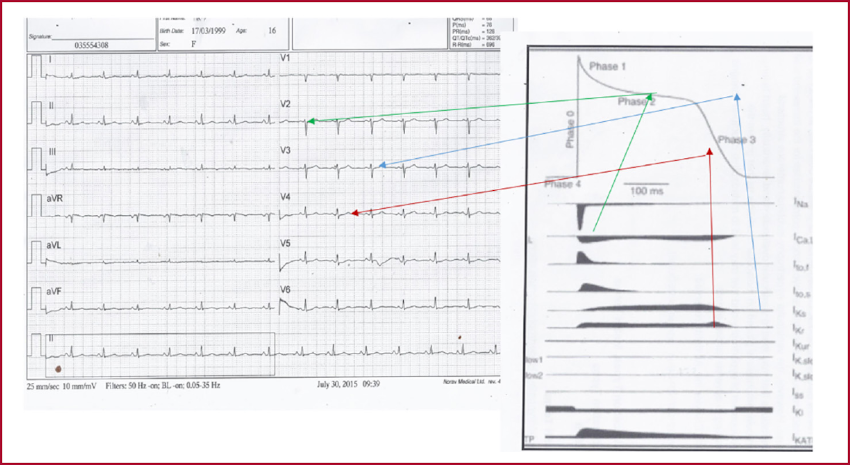
The red arrow points to the relationship between the fast rectifier potassium current and phase 3 of the action potential (AP), which conditions the slow and attenuated descending limb of the T wave (with respect to the male T wave).
The blue arrow points to the ratio of the slow rectifier potassium current with the junction of phases 2 and 3 of the AP, which forms the peak of the attenuated T wave.
The green arrow points to the relationship between the calcium current, phase 2 of the AP, and the long, attenuated ST segment.
From Nerbonne JM, Kass RS. Molecular physiology of cardiac repolarization. Physiol Rev. 2005;85:1205-53. Modified by Sclarovsky S
REFERENCES
1. Ahuja P, Sdek P, MacLellan WR. Cardiac myocyte cell cycle control in development, disease, and regeneration. Physiol Rev 2007;87:521-44. https://doi.org/10.1152/physrev.00032.2006 .
2. Cysarz D, Linhard M, Edelhäuser F, Längler A, Van Leeuwen P, Henze G, et al. Symbolic patterns of heart rate dynamics reflect cardiac autonomic changes during childhood and adolescence. Auton Neurosci 2013;178:37-43. https://doi.org/10.1016/j.autneu.2013.02.003 .
3. Hu A, Jiao X, Gao E, Koch WJ, Sharifi-Azad S, Grunwald Z, Ma XL, Sun JZ. Chronic beta-adrenergic receptor stimulation induces cardiac apoptosis and aggravates myocardial ischemia/reperfusion injury by provoking inducible nitric-oxide synthase-mediated nitrative stress. J Pharmacol Exp Ther 2006;318:469-75. https://doi.org/10.1124/jpet.106.102160 .
4. Lohse MJ, Benovic JL, Codina J, Caron MG, Lefkowitz RJ. beta-Arrestin: a protein that regulates beta-adrenergic receptor function. Science 1990;248:1547-50. https://doi.org/10.1126/science.2163110 .
5. Cotecchia S, Del Vescovo CD, Colella M, Caso S, Diviani D. The alpha1-adrenergic receptors in cardiac hypertrophy: signaling mechanisms and functional implications. Cell Signal 2015;27:1984-93. https://doi.org/10.1016/j.cellsig.2015.06.009 .
7. Kelly D, Mackenzie L, Hunter P, Smaill B, Saint DA. Gene expression of stretch-activated channels and mechanoelectric feedback in the heart. Clin Exp Pharmacol Physiol 2006;33:642-8. https://doi.org/10.1111/j.1440-1681.2006.04392.x .
8. Stones R, Gilbert SH, Benoist D, White E. Inhomogeneity in the response to mechanical stimulation: cardiac muscle function and gene expression. Prog Biophys Mol Biol 2008;97:268-81. https://doi.org/10.1016/j.pbiomolbio.2008.02.010 .
9. Esposito G, Rapacciuolo A, Naga Prasad SV, Takaoka H, Thomas SA, Koch WJ , et al . Genetic alterations that inhibit in vivo pressure-overload hypertrophy prevent cardiac dysfunction despite increased wall stress. Circulation 2002;105:85-92. https://doi.org/10.1161/hc0102.101365 .
10. Trombitás K, Granzier H. Actin removal from cardiac myocytes shows that near Z line titin attaches to actin while under tension. Am J Physiol 1997;273:C662-70. https://doi.org/10.1152/ajpcell.1997.273.2.C662 .
11. Taegtmeyer H, Sen S, Vela D. Return to the fetal gene program: a suggested metabolic link to gene expression in the heart. Ann N Y Acad Sci 2010;1188:191-8. https://doi.org/10.1111/j.1749-6632.2009.05100.x .
12. El Gebeily G, El Khoury N, Mathieu S, Brouillette J, Fiset C. Estrogen regulation of the transient outward K(+) current involves estrogen receptor α in mouse heart. J Mol Cell Cardiol 2015;86:85-94. https://doi.org/10.1016/j.yjmcc.2015.07.013 .
13. Zhu Y, Ai X, Oster RA, Bers DM, Pogwizd SM. Sex differences in repolarization and slow delayed rectifier potassium current and their regulation by sympathetic stimulation in rabbits. Pflugers Arch 2013;465:805-18. https://doi.org/10.1007/s00424-012-1193-9 .
14. Chu SH, Goldspink P, Kowalski J, Beck J, Schwertz DW. Effect of estrogen on calcium-handling proteins, beta-adrenergic receptors, and function in rat heart. Life Sci 2006;79:1257-67. https://doi.org/10.1016/j.lfs.2006.03.037 .
15. Liu XK, Katchman A, Whitfield BH, Wan G, Janowski EM, Woosley RL, et al. In vivo androgen treatment shortens the QT interval and increases the densities of inward and delayed rectifier potassium currents in orchiectomized male rabbits. Cardiovasc Res 2003;57:28-36. https://doi.org/10.1016/s0008-6363(02)00673-9 .
16. Michels G, Er F, Eicks M, Herzig S, Hoppe UC. Long-term and immediate effect of testosterone on single T-type calcium channel in neonatal rat cardiomyocytes. Endocrinology 2006;147:5160-9. https://doi.org/10.1210/en.2006-0186 .